Sơ đồ tư duy Sinh học 12 chương 1 ngắn gọn, dễ hiểu | Myphamthucuc.vn
Nội dung bài viết
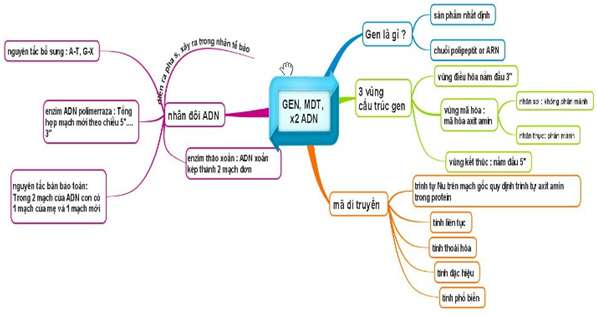
TÓM TẮT LÝ THUYẾT CHƯƠNG I: GEN, MÃ DI TRUYỀN
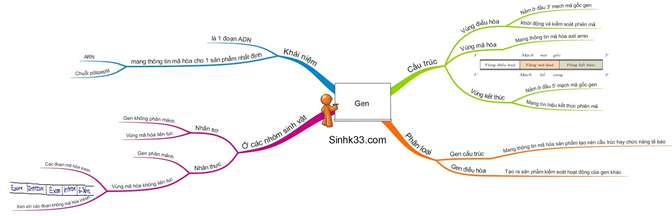
I. Gen
1. Khái niệm
– Gen là một đoạn ADN mang thông tin mã hoá cho một chuỗi pôlipeptit hay một phân tử ARN.
– Vd: Gen Hba mã hoá chuỗi pôlipeptit a, gen tARN mã hoá cho phân tử tARN.
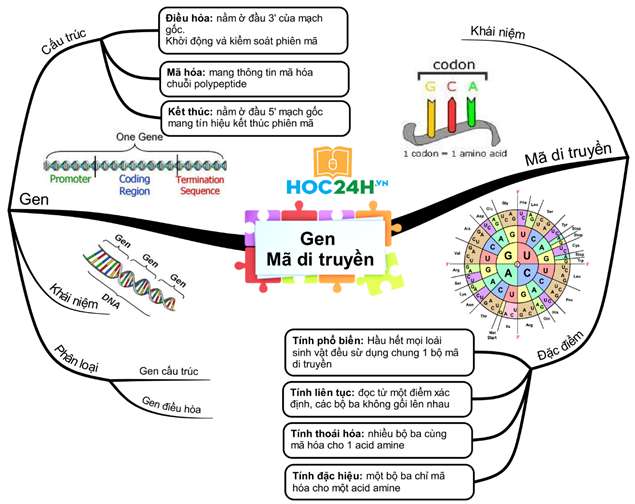
2. Cấu trúc chung của gen cấu trúc (gen mã hóa chuỗi Polipepetit)
|
Vùng điều hòa |
Vùng mã hóa |
Vùng kết thúc |
|
| Vị trí | Nằm ở đầu 3’ của mạch mã gốc. | Nằm ở giữa gen. | Nằm ở đầu 5’ của mạch mã gốc. |
| Chức năng | Có trình tự nuclêôtit đặc biệt giúp ARN pôlymeraza có thể nhận biết, liên kết để khởi động và điều hòa quá trình phiên mã. |
Mang thông tin mã hóa các axit amin. – SV nhân sơ: gen không phân mảnh (chỉ có exon) – SV nhân thực: gen phân mảnh (đoạn exon xen kẽ đoạn intron) |
Mang tín hiệu kết thúc phiên mã. |
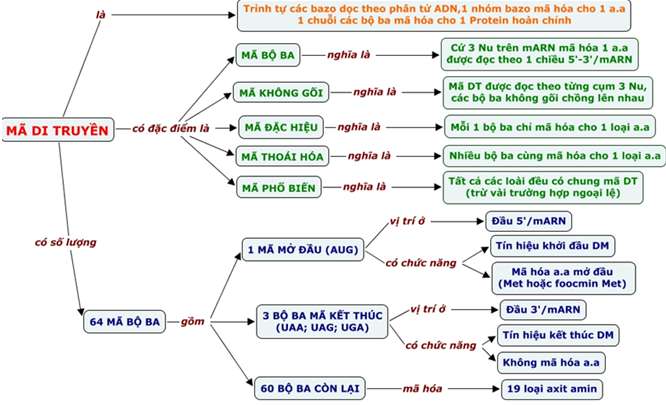
II. Mã di truyền
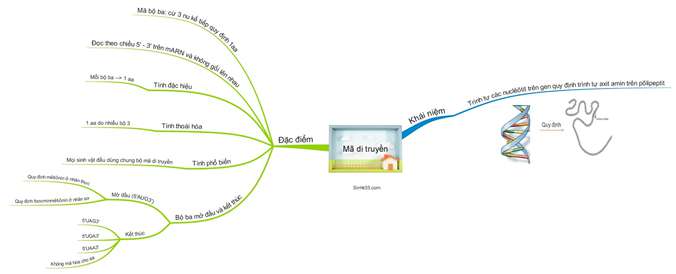
1. Khái niệm
– Mã di truyền là trình tự sắp xếp các nuclêôtit trong gen (mạch gốc) quy định trình tự sắp xếp các axit amin trong prôtêin.
– Mã di truyền (côđon) là mã bộ ba (ba nuclêôtit mã hóa một axit amin). Trong ADN có 4 loại nuclêôtit à có tất cả 43= 64 bộ ba, nhưng chỉ có 61 bộ ba tham gia mã hóa axit amin (trừ 3 bộ ba: UAA, UAG, UGA là 3 bộ ba kết thúc không mã hóa axit amin).
– Bộ ba AUG là mã mở đầu với chức năng khởi đầu dịch mã và mã hóa axit amin mêtiônin (ở sinh vật nhân thực) hay mã hóa axit amin foocmin mêtiônin (ở sinh vật nhân sơ).
2. Đặc điểm
(1) Mã di truyền được đọc từ một điểm theo chiều 3’ → 5’, theo từng bộ ba, không gối lên nhau.
(2) Mã di truyền có tính phổ biến (tất cả các loài có chung một bộ mã di truyền, trừ một vài ngoại lệ)
(3) Mã di truyền có tính đặc hiệu: 1 bộ ba mã hóa 1 a.amin
(4) Mã di truyền có tính thoái hoá: 1 aa. được mã hóa từ nhiều bộ ba khác nhau.
III. Quá trình nhân đôi ADN
1. Thời điểm: Quá trình nhân đôi ADN tại pha S của kì trung gian.
2. Nơi diễn ra: Nhân tế bào (TB nhân thực); vùng nhân (TB nhân sơ)
3. Chiều tổng hợp: 5’ – 3’
4. Diễn biến
(1) Bước 1:(Tháo xoắn phân tử ADN)
-Nhờ các enzim tháo xoắn, 2 mạch phân tử ADN tách nhau dần → chạc hình chữ Y và lộ 2 mạch khuôn.
(2) Bước 2:(Tổng hợp các mạch ADN mới)
– Enzim ADN pôlimeraza tổng hợp 2 mạch mới nhờ 2 mạch khuôn theo nguyên tắc bổ sung (A=T; G≡X):
+ Mạch tổng hợp liên tục: Có mạch khuôn là chiều 3’→ 5’
+ Mạch tổng hợp ngắt quãng: Có mạch khuôn là chiều 5’→ 3’. Chúng tổng hợp theo từng đoạn (Okazaki) rồi được nối lại với nhau nhờ enzim nối (ligaza).
(3) Bước 3:( 2 phân tử ADN được tạo thành)
– Mỗi phân tử ADN mới gồm 2 mạch:
+ 1 mạch của phân tử ADN ban đầu (bán bảo toàn)
+ 1 mạch mới được tổng hợp.
5. Kết quả: một phn tử ADN qua n lần nhân đôi tạo thành 2n phân tử ADN con
6. Mục đích:
– Tạo ra 2 crômatit dính nhau trong NST đảm bảo NST tự nhân đôi (phân chia tế bào).
– Đảm bảo giữ nguyên về cấu trúc và hàm lượng ADN qua các thế hệ.
– Ổn định các đặc điểm của loài từ thế hệ này sang thế hệ khác.
IV. Phiên mã
1. Cấu trúc và chức năng của các loại ARN:
|
|
mARN (ARN thông tin) |
tARN (ARN vận chuyển) |
rARN (ARN ribôxôm) |
| Cấu tạo | – Là một mạch đơn thẳng, có 600-1500 đơn phân gọi là ribônuclêôtit (rNu).- Có 4 loại ribônuclêôtit: A, U, G, X.- Đầu 5’ có trình tự nuclêôtit đặc hiệu.- Liên kết cộng hóa trị giữa các ribônuclêôtit, không có kiên kết hidro. | – Là một mạch đơn tự xoắn, có 80–100 đơn phân là ribônuclêôtit (rNu).- Có 4 loại ribônuclêôtit: A, U, G, X.- Một đầu mút gắn với axit amin, đầu kia tự do.- Liên kết cộng hóa trị và liên kết hidro theo nguyên tắc bổ sung giữa các ribônuclêôtit.- Có 1 bộ ba đối mã đặc hiệu (anticôđon). | – Là một mạch đơn tự xoắn, gồm 2 tiểu đơn vị tồn tại riêng rẽ trong tế bào chất. Khi tổng hợp, chúng liên kết với nhau thành ribôxôm hoạt động chức năng.- Có 70% ribônuclêôtit có liên kết hidro nguyên tắc bổ sung. |
| Chức năng | – Là bản sao mã, mang thông tin di truyền từ trong nhân ra ngoài tế bào chất.- Làm khuôn để dịch mã tổng hợp nên chuỗi pôlypeptit. | – Mang axit amin tới ribôxôm tham gia dịch mã trên mARN thành trình tự các axit amin trên chuỗi pôlypeptit.- tARN có thể sử dụng nhiều lần, qua nhiều thế hệ tế bào. | – Kết hợp với prôtêin tạo nên ribôxôm – nơi tổng hợp chuỗi pôlypeptit. |
2. Cơ chế phiên mã: (Tổng hợp ARN)
a. Phiên mã là quá trình tổng hợp ARN trên mạch khuôn ADN.
b. Diễn biến
(1) ARN pôlimeraza bám vào vùng điều hòa làm gen tháo xoắn lộ mạch gốc có chiều 3’→5’ bắt đầu phiên mã. ARN polimeraza trượt trên mạch gốc theo chiều 3’→5’.
(2) mARN được tổng hợp theo chiều 5’→ 3’, mỗi nu trên mạch gốc liên kết với nu tự do theo nguyên tắc bổ sung A-U, G- X, T-A, X-G (vùng nào trên gen được phiên mã song thì sẽ đóng xoắn ngay).
(3) Khi ARN pôlimeraza gặp tín hiệu kết thúc thì dừng phiên mã. Một phân tử mARN được giải phóng.
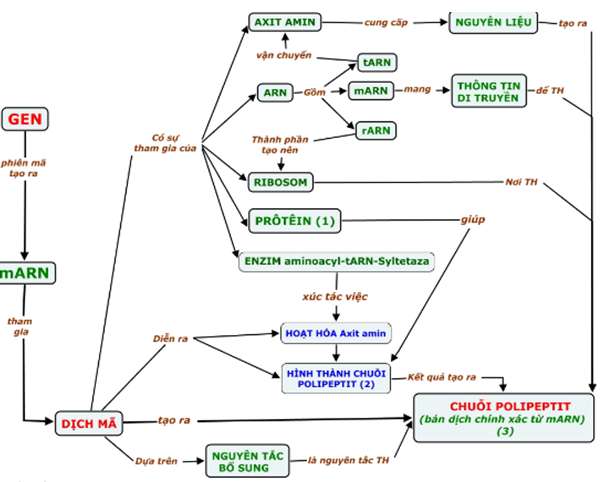
c. Kết quả: Tạo nên phân tử mARN mang thông tin di truyền từ gen tới ribôxôm để làm khuôn trong tổng hợp prôtêin.
*** Lưu ý: Ở sinh vật nhân thực mARN sau khi tổng hợp sẽ cắt bỏ các đoạn Intron, nối các đoạn Exon tạo thành mARN trưởng thành sẵn sằng tham gia dịch mã.
V. Dịch mã
1. Hoạt hóa axit amin: Trong tế bào chất, nhờ các enzim đặc hiệu và năng lượng ATP, mỗi axit amin được hoạt hóa và gắn với tARN tương ứng tạo nên phức hợp axit amin tARN (aa–tARN).
2. Tổng hợp chuỗi pôlypeptit:
a. Mở đầu:
– Tiểu đơn vị bé của ribôxôm gắn với mARN ở vị trí nhận biết đặc hiệu.
– Bộ ba đối mã của phức hợp mở đầu (aamở đầu – tARN) Met – tARN (UAX) bổ sung chính xác với côđon mở đầu (AUG) trên mARN.
– Tiểu đơn vị lớn của ribôxôm tiến vào gắn với tiểu đơn vị bé tạo ribôxôm hoàn chỉnh, sẵn sàng tổng hợp chuỗi pôlipeptit.
b. Kéo dài chuỗi pôlypeptit:
– aa1 – tARN gắn bổ sung với côđon thứ 2 trên mARN theo nguyên tắc bổ sung (A=U, GºX và U=A, XºG).
– Liên kết peptit hình thành giữa aamở đầu và aa1.
– Ribôxôm dịch chuyển đi một côđon trên mARN, aa2–tARN gắn bổ sung với côđon thứ 3 trên mARN, aa2 liên kết aa1 bằng liên kết peptit.
– Ribôxôm trượt một côđon trên mARN và cứ tiếp tục cho đến cuối mARN.
c. Kết thúc: Khi ribôxôm tiếp xúc với 1 trong 3 mã kết thúc trên mARN (UAA, UAG, UGA) thì quá trình dịch mã hoàn tất.
*** Lưu ý: Trong quá trình dịch mã, mARN thường không gắn với từng ribôxôm riêng rẽ mà đồng thời gắn với một nhóm ribôxôm gọi là pôliribôxôm (gọi tắt là pôlixôm) giúp tăng hiệu suất tổng hợp prôtêin.
VI. Điều hòa hoạt động gen
1. Khái niệm: Điều hòa hoạt động gen là điều hòa lượng sản phẩm do gen tạo ra. Tế bào chỉ tổng hợp prôtêin cần thiết vào lúc thích hợp với lượng cần thiết. Ở sinh vật nhân sơ: chủ yếu điều hòa ở mức phiên mã (điều hòa lượng mARN tổng hợp trong tế bào).
2. Cấu trúc của Opêron Lac: gồm 3 vùng
(1) Vùng khởi động (Promoter): nơi enzim ARN-pôlymeraza bám vào và khởi đầu phiên mã.
(2) Vùng vận hành (Operator): trình tự nuclêôtit đặc biệt, nơi gắn của prôtêin ức chế để ngăn cản phiên mã.
(3) Nhóm gen cấu trúc (Z, Y, A): tổng hợp enzim phân giải đường lactôzơ để cung cấp năng lượng cho tế bào.
* Chức năng của gen điều hòa R: Gen điều hòa R khi hoạt động sẽ tổng hợp nên prôtêin ức chế. Prôtêin này có khả năng liên kết với vùng vận hành (O) dẫn đến ngăn cản quá trình phiên mã (gen đều hịa khơng nằm trong Opron)
3. Điều hòa hoạt động của Opêron Lac:
a. Khi môi trường không có lactôzơ: Gen điều hòa tổng hợp prôtêin ức chế. Prôtêin này liên kết với vùng vận hành (O) ngăn cản quá trình phiên mã làm cho các gen cấu trúc không hoạt động.
b. Khi môi trường có lactôzơ:
– Một số phần tử lactôzơ liên kết với prôtêin ức chế làm biến đổi cấu hình không gian ba chiều của nó → prôtêin ức chế không thể liên kết được với vùng vận hành (O) nên ARN pôlymeraza có thể liên kết được với vùng khởi động (P) để tiến hành phiên mã.
– Sau đó, các phân tử của mARN của các gen cấu trúc Z, Y, A được dịch mã tạo ra các enzim phân giải đường lactôzơ. Khi đường lactôzơ bị phân giải hết thì prôtêin ức chế lại liên kết với vùng vận hành (O) và quá trình phiên mã bị dừng lại.
VII. Đột biến gen
1. Khái niệm
– Đột biến gen là những biến đổi trong cấu trúc của gen, liên quan đến một cặp nuclêôtit (đột biến điểm) làm thay đổi trình tự nu tạo ra alen mới.
– Tất cả các gen có thể bị đột biến với tần số thấp (10-6 – 10-4)
– Thể đột biến là cá thể mang gen đột biến đã biểu hiện ra kiểu hình
2. Các dạng đột biến gen
(1) Đột biến thay thế một cặp nuclêôtit
(2) Đột biến thêm hoặc mất một cặp nuclêôtit: Mã di truyền bi đọc sai kể từ vị trí xảy ra đột biến → thay đổi trình tự axit amin → thay đổi chức năng protein.
3. Nguyên nhân
– Bên ngoài: do tác nhân vật lý (tia phóng xạ, tia tử ngoại…), hoá học (các hoá chất 5BU, NMS…) hay sinh học(1 số virut…).
– Bên trong: do rối loạn các quá trình sinh lí hóa sinh trong tế bào.
4. Cơ chế phát sinh đột biến gen
(1) Sự kết cặp không đúng trong nhân đôi ADN
– Trong quá trình nhân đôi, sự kết cặp không theo nguyên tắc bổ sung → phát sinh đột biến gen.
Ví dụ: G* (dạng hiếm) kết hợp T: Tạo đột biến G – X thành A – T
(2) Tác động của các tác nhân gây đột biến
– Tia tử ngoại (UV): làm 2 bazơ Timin trên cùng 1 mạch liên kết với nhau → đột biến gen.
– 5-brôm uraxin (5BU) gây đột biến thay thế cặp A-T bằng G-X
A – T → A – 5BU → G – 5BU → G – X
– Virut viêm gan B, virut hecpet…→ đột biến.
5. Hậu quả
(1) Đột biến gen có thể có hại, có lợi, vô hại.
(2) Phần lớn đột biến điểm thường vô hại (trung tính)
(3) Tính có hại của đột biến phụ thuộc môi trường, tổ hợp gen.
6. Vai trò và ý nghĩa của đột biến gen
a. Đối với tiến hoá
– Đột biến gen làm xuất hiện các alen mới tạo ra biến dị di truyền phong phú là nguồn nguyên liệu cho tiến hoá.
b. Đối với thực tiễn
– Cung cấp nguyên liệu cho quá trình tạo giống và trong nghiên cứu di truyền.
7. Biểu hiện:
a. Đột biến sôma (đột biến sinh dưỡng):
– Xảy ra trong nguyên phân của tế bào sinh dưỡng rồi được nhân lên trong 1 mô. Nếu là đột biến trội nó sẽ biểu hiện ở một phần cơ thể tạo nên thể khảm.
– Đột biến sôma có thể nhân lên bằng sinh sản sinh dưỡng nhưng không thể di truyền qua sinh sản hữu tính.
b. Đột biến giao tử (đột biến sinh dục):
– Xảy ra trong giảm phân của tế bào sinh dục, qua thụ tinh đi vào hợp tử. Nếu đột biến trội sẽ biểu hiện ngay ở thế hệ sau, nếu đột biến lặn sẽ đi vào hợp tử trong cặp gen dị hợp và tồn tại trong quần thể bị gen trội lấn át, qua giao phối đột biến lặn lan truyền trong quần thể và hình thành tổ hợp đồng hợp lặn mới biểu hiện thành thể đột biến.
– Di truyền cho thế hệ sau bằng sinh sản hữu tính.
c. Đột biến tiền phôi:
– Xảy ra ở những lần nguyên phân đầu tiên của hợp tử (trong giai đoạn 2 – 8 phôi bào). Nhờ nguyên phân sẽ nhân lên và biểu hiện trong toàn bộ cơ thể.
– Di truyền cho thế hệ sau bằng sinh sản hữu tính.
VIII. Nhiễm sắc thể
1. Hình thái NST: gồm 3 phần:
(1) Tâm động: là vị trí liên kết với thoi phân bào giúp NST di chuyển về các cực của tế bào trong quá trình phân bào.
(2) Vùng đầu mút: có tác dụng bảo vệ các NST và làm cho các NST không dính vào nhau.
(3) Các trình tự khởi đầu nhân đôi: là những điểm mà ở đó, ADN bắt đầu nhân đôi.
2. Đặc điểm:
– NST có hình dạng và kích thước đặc trưng cho loài, nhìn rõ nhất ở kì giữa của nguyên phân khi chúng co xoắn cực đại.
– Ở sinh vật nhân thực: mỗi tế bào chứa từng phân tử ADN liên kết với các loại prôtêin khác nhau (chủ yếu loại histôn).
– Ở sinh vật nhân sơ: mỗi tế bào chỉ chứa một phân tử ADN mạch kép, có dạng vòng và chưa có cấu trúc NST.
3. Cấu trúc hiển vi:
– Mỗi nuclêôxôm gồm 8 phân tử prôtêin (chủ yếu dạng histôn) được quấn quanh vòng xoắn (khoảng 146 cặp nuclêôtit).
– Có 3 mức độ xoắn khác nhau:
+ Mức xoắn 1: chuỗi nuclêôxôm (sợi cơ bản) có đường kính 11nm.
+ Mức xoắn 2: sợi chất nhiễm sắc có đường kính 30nm.
+ Mức xoắn 3: siêu xoắn (vùng xếp cuộn) có đường kính 300nm tạo thành crômatit có đường kính 700nm.
(Đoạn ADN (146 cặp Nu) + 8 prôtêin Histôn → Nuclêôxôm → Sợi cơ bản (đk 11nm) → Sợi chất nhiễm sắc (đk 30nm) → Sợi siêu xoắn (300nm) → Crômatic (đk 700nm))
IX. Đột biến cấu trúc nhiễm sắc thể
1. Khái niệm: Đột biến cấu trúc NST là những biến đổi trong cấu trúc NST, bản chất là sự sắp xếp lại các gen trên NST.
2. Nguyên nhân:
– Môi trường: do các tác nhân vật lí trong ngoại cảnh như: tia phóng xạ, tia tử ngoại, sốc nhiệt hay các hóa chất.
– Trong cơ thể: rối loạn quá trình sinh lí, sinh hóa, trao đổi chất trong tế bào và cơ thể.
3. Các dạng:
a. Mất đoạn
– NST bị mất 1 đoạn làm giảm số lượng gen trên NST → thường gây chết.
Ví dụ: Mất đoạn NST số 5 gây hội chứng mèo kêu
– Ưng dụng: loại khỏi NST những gen không mong muốn ở 1 số giống cây trồng.
b. Lặp đoạn
– Một đoạn NST được lặp lại một hay nhiều lần → tăng số lượng gen trên NST.
– Làm tăng hoặc giảm cường độ biểu hiện của tính trạng (có lợi hoặc có hại).
Ví dụ: Lúa Đại mạch đột biến lặp đoạn → tăng hoạt tính enzim amilaza.
c. Đảo đoạn:
– Một đoạn NST bị đứt ra rồi đảo ngược 1800 và nối lại → làm thay đổi trình tự gen trên NST → làm ảnh hưởng đến hoạt động của gen.
Ví dụ: ở muỗi, đột biến đảo đoạn lặp lại nhiều lần → tạo nên loài mới.
4. Chuyển đoạn:
– Sự trao đổi đoạn NST xảy ra giữa các NST không tương đồng (Chú thích hình bên) → thay đổi kích thước, cấu trúc gen, nhóm gen liên kết → giảm khả năng sinh sản.
Ví dụ: ở người, đột biến chuyển đoạn giữa NST số 22 và NST số 9 → NST 22 ngắn hơn → ung thư máu ác tính.
5. Cơ chế phát sinh:
– Các tác nhân gây đột biến trong ngoại cảnh hoặc trong tế bào làm cho NST bị đứt, gãy ảnh hưởng tới quá trình tự nhân đôi của NST, trao đổi chéo của các crômatit.
– Dạng đột biến này thực chất là sự sắp xếp các khối gen trên NST và giữa các NST. Do vậy có thể làm thay đổi hình dạng và cấu trúc của NST.
X. Đột biến số lượng nhiễm sắc thể
Khái niệm: Đột biến số lượng NST là sự thay đổi số lượng NST trong tế bào.
Gồm 2 loại: đột biến lệch bội ( dị bội ), đột biến đa bội
1. Đột biến lệch bội
a. Khái niệm:
Đột biến dị bội làm thay đổi số lượng NST ở 1 hay 1 số cặp tương đồng.
b. Phân loại
-Thể mộ (2n-1): 1 cặp NST mất 1 NST
-Thể không (2n -2) : 1 cặp NST mất 2 NST
-Thể ba (2n +1): 1 cặp NST thêm 1 NST
-Thể bốn (2n +4): 1 cặp NST thêm 2 NST
c. Cơ chế phát sinh
*) Trong giảm phân
– Do sự phân ly NST không bình thường ở 1 hay 1 số cặp kết quả tạo ra các giao tử thiếu, thừa NST (n -1; n + 1 giao tử lệch nhiễm).
– Các giao tử này kết hợp với giao tử bình thường ® thể lệch bội.
*) Trong nguyên phân
– Trong nguyên phân một số cặp NST phân ly không bình thường hình thành tế bào lệch bội. Tế bào lệch bội tiếp tục nguyên phân ® 1 phần cơ thể có các tế bào bị lệch bội ® thể khảm.
d. Hậu quả: tử vong, giảm sức sống, giảm khả năng sinh sản…
e. Ý nghĩa: Đột biến lệch bội cung cấp nguyên liệu cho tiến hoá và chọn giống.
2. Đột biến đa bội
a. Khái niệm
– Là dạng đột biến làm tăng 1 số nguyên lần bộ NST đơn bội của loài và lớn hơn 2n
– Phân loại:
+ Theo bộ NST: đa bội lẻ (3n, 5n, 7n…); đa bội chẵn (4n, 6n, 8n,…)
+ Theo nguồn gốc: tự đa bội (sự gia tăng số bộ NST từ 1 loài); dị đa bội sự gia tăng số bộ NST từ 2 loài khác nhau)
b. Cơ chế phát sinh
* Tự đa bội
– Dạng 3n là do sự kết hợp giữa giao tử n với giao tử 2n (giao tử lưỡng bội).
– Dạng 4n là do sự kết hợp giữa 2 giao tử 2n hoặc trong lần nguyên phân đầu tiên của hợp tử tất cả các cặp NST không phân ly.
* Dị đa bội
– Do hiện tượng lai xa và đa bội hoá.
c. Hậu quả và vai trò của đột biến đa bội
– Tế bào đa bội thường có số lượng ADN tăng gấp bội → tế bào to, cơ quan sinh dưỡng lớn, sinh trưởng phát triển mạnh khả năng chống chịu tốt. . .
– Đột biến đa bội đóng vai trò quan trọng trong tiến hoá (hình thành loài mới) và trong trồng trọt (tạo cây trồng năng suất cao. . . )
SƠ ĐỒ TƯ DUY CHƯƠNG I: GEN, MÃ DI TRUYỀN
MẪU SỐ 1
MẪU SỐ 2
MẪU SỐ 3
MẪU SỐ 4
MẪU SỐ 5
MẪU SỐ 6
Có thể bạn quan tâm










Bài viết hay nhất
Các Kiểm Tra Hạn Sử Dụng Của Kem Chống Nắng Skin Aqua Nhật Bản

Câu hỏi ôn tập bài Sau phút chia li chọn lọc chi tiết

Bài thơ “Ngắm trăng” được sáng tác trong hoàn cảnh nào?

Phân tích tâm trạng nhân vật Liên khi chờ tàu năm 2023

Kem Dưỡng Estee Lauder Thật Giả Qua Các Dung Tích, So Sánh Và Nhận Biết Serum Advance Night Repair

Giới thiệu về Ngô Gia văn phái năm 2023

Nhân vật Mị sau khi về làm dâu nhà thống lí Pá Tra trong Vợ chồng A Phủ

Phân tích bức tranh tứ bình trong bài Nhớ rừng của Thế Lữ năm 2023

Viết thư cho Cô giáo cũ nhân ngày 20/11 năm 2023

Soạn bài Vừa nhắm mắt vừa mở cửa sổ
Chủ đề của truyện Tấm Cám là gì

Cách Vẽ Mặt Nạ Hình Con Vật Hươu Cho Bé, Cách Làm Mặt Nạ Hình Con Thỏ Chơi Trung Thu

Cách Dùng Kem Dưỡng Da Johnson Baby Nắp Xanh Có Tác Dụng Gì? Sử Dụng Ra Sao?

Câu hỏi bài Hai đứa trẻ chọn lọc